
Compaction Factor Test – Workability of Concrete
Compaction Factor Test is designed in such a way that it can be used only in laboratory but in some cases, it can be used for field concrete tests. The compacting factor test has been developed at the Road Research Laboratory in United Kingdom. This test is one of the most accurate test performed in order to determine the workability of concrete.
This test works on the principle of determining the degree of compaction achieved by a standard amount of work done by allowing the concrete to fall through a standard height. The degree of compaction, called the compacting factor is measured by the density ratio i.e., the ratio of the density actually achieved in the test to density of same concrete fully compacted.
The apparatus of compaction factor test is shown below.

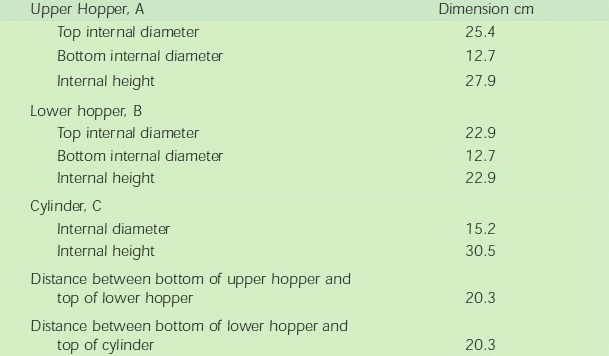
Dimension of the Test Apparatus is given below.
Procedure of Compacting Factor test:
- Prepare a concrete mix in the ratio of 1:2:4
- With the help of a trowel, fill the freshly prepared concrete in the top upper of the apparatus. The concrete should be filled to the brim of the hopper and level it of with trowel.
- Now open the trap of the upper hopper, so that the concrete falls in the lower hopper.
- After all concrete falls from the upper hopper to lower one. Then again open the trap of the lower hopper. Let the concrete falls on the cylinder.
- Now take the weight of the cylinder in which concrete had felled. Let this weight be “The weight of partially compacted concrete (W1)“.
- Empty the cylinder.
- Now again, fill concrete in the cylinder in three layers with 25 blows for each layer using tamping rod. Fill concrete to the top of cylinder and scrape excess concrete above the brim.
- Now take the weight of the cylinder in which concrete we filled. Let this weight be “The weight of fully compacted concrete (W2)“.
The compacting factor of concrete can be found out using the formula,
= (Weight of Partially Compacted Concrete W1)/(Weight of Partially Compacted Concrete W2)
Compacting Factor test Or Slump Cone Test. Which is Best?
- This test is more accurate and sensitive than the slump cone test.
- In this test, Concrete mixes of very low workability can be tested out. Where in slump test it is difficult and it gives inaccurate results for dry mix.